The Yawn of Time
Of N-dimensional hypercubes and a calculated enzyme rate
I spent nearly all of the last week in my office. Sitting around waiting for something to happen; there is only so much editing and e-mail a person can do. Still one has to remember a scientist’s life is not all bubbling green liquids and shouts of Eureka! Or Bakersfield! (look at a map of California). There is planning and plotting, plotting and planning. Especially to start new things. The carpenter’s adage “measure twice, cut once” applies here, with measurement involving an N-dimensional hypercube.
This measuring involved talking things through with various folk and ordering some supplies. I ordered two “Really Useful Boxes” (yes, that is their brand name) that are large and solid black, with a tight fitting lid. I myself ordered special scotch darkroom tape, the kind that filled up my Ph. D. work and that is awesome for keeping things light-tight. Hilariously, this had to be ordered from USA, cost as much for the tape as for shipping, but so be it. Both of these items are needed for the dark project. This is the one that does not involve confocal or lab weed, and needs to have plants grown and experiments conducted in complete darkness. I get to be in the dark literally, but I hope not figuratively.
The small amount of lab work I did accomplish got off to an embarrassing start. The week before, the helpful Kamal had shown me my 5 foot of bench space and, on Monday, I thought I ought to colonize it. For one thing, I had a package of maize seed that didn’t belong on my desk, where it might receive the losing end of a coffee spill. I also had a brand new, whiter-than-white, lab coat. Yes, I’ll keep the safety police sweet; when in Rome and all that (see blog post 1); maybe I should call it a toga? Thus properly attired, I got into the lab and could not remember which bit of bench was mine. Ummm… The lab is large with row after row of benches, all looking more or less the same. I cruised around hoping for the flash of recognition, in vain.
Feeling ridiculous, I went back into the toga room interposed between lab and offices, a kind antechamber with 1000 togas on pegs, with each peg bearing its owners name on a bright piece of yellow tape. None had my name; none were nameless; no place to hang my toga. Could it get more ridiculous than this? Later in the week, Morag found an unnamed peg, hiding in the corner, which I emblazoned with my name, and she pointed out my bench, which I marked with a strip of orange label tape on the shelf.
Best of all, on Friday, I plated some seed. Our good friend, lab weed (see blog post 2) has tiny seed, more or less the same size as a poppy seed, or perhaps the size of the stop at the end of this sentence. Plating seeds describes putting the seed on a Petri plate that contains agar medium. The agar provides a firm surface, rather like Jell-o and about as tasty. The medium contains nutrients (good old nitrogen, potassium, phosphorus) to build healthy plants. The seeds are placed on the surface of the agar, in the Petri plate, and the plates are stood on their sides. After germination, the plant grows well on the surface of the agar. It is amusing to use Petri plates to grow seedlings in, as if they were some glorified bacterium.
Plating seed has to solve two problems. Because the seeds are so small, you cannot just pick them up with your fingers. Because the agar-medium is nutritious, bacteria and fungi eagerly grow there too, especially when the medium contains sucrose, which it often does. Even though it seems strange, the little lab weed seedlings have a sweet root (I mean they enjoy medium with sucrose). To exclude members of these other kingdoms, it is typical to plate seeds in a laminar flow hood, basically a clean bench. And to pick up the wee things, scientists have come up with a host of tricks. My favorite and happily one that seems in vogue here, is to pick up a dozen or so seed with a pipette (figure). The seeds are spread out in 20 µL within the pipette tip and by slowly squeezing one can deliver them one by one. Sometimes two seed come out at the same time but that is ok.

I used to plate lab weed seed all the time, but I haven’t done for four or five years, maybe longer. I was chuffed to see that my skill remained, evidently like riding a bicycle. I put the plates in the fridge for a couple of days to give the seeds a cold exposure, which they require to germinate. I’ll put them in the growth chamber on Tuesday. When the roots are a few cm long, I’ll take some more movies on the confocal.
OK, I am going to end with the story of a calculation. Talking about this is perhaps unfair, it relates to a project leftover from my past life in Massachusetts, and talking about it is certainly geeky. But as I spent a whole afternoon on it, seems fair game. Furthermore, this kind of reasoning exemplifies how that N-dimensional hypercube has to be measured before the tubes of phosphorescent green fluid can be boiled.
I want to know how fast a particular enzyme cranks out its reaction product. In general terms, an enzyme is (usually) a protein that catalyzes a chemical reaction. The words chemical reaction encompass a wide diversity of changes; nevertheless, the typical reaction for an enzyme is to convert one molecule to another. And this is the kind of reaction that concerns me. The original molecule is called the substrate (what the enzyme acts on) and the molecule that results is called the product (what the enzyme produces). To make the reaction happen, the enzyme has to bind the substrate; wiggle to encourage the chemical bonds in the substrate to rearrange, thus forming the product; and then release the product. These steps will take a defined amount of time that will depend on the players (enzyme, substrate, product), environment (temperature, concentrations), and the required wiggles. Therefore, the rate at which enzymes work varies hugely, with the most slothful catalyzing a few reactions per second and the busiest doing 10 to the twelfth reactions per second or maybe even more. This means that for any given enzyme of interest, there is no way to guess its reaction rate, and instead it must be measured.
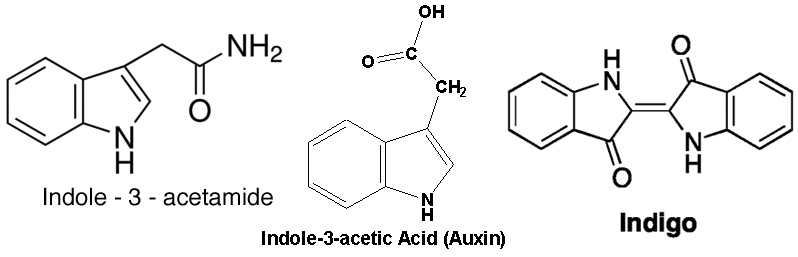
The enzyme of interest to me rejoices under the name of indole-3-acetamide hydrolase, aka IAAH because it is a bit of a mouthful even for scientists. The suffix “-ase” is widely used to name the reaction of enzymes, which in this case is a hydrolysis, hence hydrolase. The indole-3-acetamide is the substrate, a small molecule that is related to the amino acid, tryptophan. The indole part also makes up the bulk of the natural dye, indigo, and the similar name is no coincidence. Now, the product of the IAAH reaction is indole-3-acetic acid (the reaction changes amide to acid), which is none other than the plant hormone, auxin. IAAH is used by several kinds of bacteria and even a few species of plant to make auxin. Chemical structures of these molecules shown below.
I have a line of lab weed in which IAAH occurs in only four cells of the root (the quiescent center). I want to know how fast those cells fill up with auxin when the cells (and hence the IAAH) are flooded with substrate. The first thing I did was to look up papers on the enzymatic properties of the enzyme. I was hoping that someone had purified the enzyme and measured it s reaction rate as a function of the number of IAAH molecules present. Then I could make some reasonable assumption about the number of IAAH molecules in those four cells and estimate how fast auxin would accumulate. Although I have not finished this literature hunt, I came up surprisingly empty.
But what I did find were measurements of IAAH reaction rate in cell extracts. That means they broke open cells and added the substrate and measured the rate at which product accumulated. This is a measure of activity of the enzyme but the problem is that they didn’t count the number of IAAH enzyme molecules in the mix so it is an activity per cell, rather than per enzyme. But then I had the idea of saying: Well ok, why not assume that the amount of IAAH present in the bacterial cell is the same as that in the quiescent center cell of the root? Since the latter quantity has to be assumed (guessed!) anyway, this is seems as good as anything else.
With that idea in mind, I had a go. First, the activity in the paper I mentioned was reported per 10 to the 9th bacterial cells. I converted to this to per mg protein by looking up how much protein a typical bacterium possesses. Then I worked out how many mg of protein a quiescent center cell might contain and their volume. Then I could use the enzyme activity per mg protein in the volume of the quiescent center and calculate how fast the cells would fill up with auxin (this of course neglects any reactions that will remove auxin). I got 40 µM in one hour, or 670 nM in one minute.
Is this reasonable? When I treat wild-type lab weed with the substrate of IAAH, nothing happens. But when I treat the special line that has IAAH in its quiescent center, they look similar to wild-type plants treated with auxin concentrations that are higher than 100 nM, and only minutes are needed for this to happen. So yes, this 670 nM number is right order of magnitude, an agreement I found immensely satisfying after a long afternoon of pushing exponents around. Of course, this agreement might be a coincidence, but still.