30/11 The cold darkness of the Universe…
“Further, the cold darkness of the Universe can be used as a renewable thermodynamic resource, even during the hottest hours of the day.” This striking quote comes from Raman et al. 2014 Nature 515: pg 540, who report on a device that sits in the sun and cools, passively (meaning without an energy source). They put it on a roof out in sunny California and it cooled to 5 degrees C below ambient. The device works by reflecting visible light almost perfectly and transmitting longish infra red (wavelengths between 8 to 13 µm) again almost perfectly. This allows more heat to pass out trough the device than is absorbed, effecting cooling. Sort of the opposite of the greenhouse effect exerted by atmospheric gasses like carbon dioxide and methane. The point of the quote is that there is not much energy in the 8 to 13 µm band and while human history could be described as finding unspoiled expanses and spoiling them, filling up this particular expanse of universal emptiness is well beyond us, even at our most rapacious. As built, the cooling power of the device was pretty decent, about 40 Watts per square meter, which is not enough to provide typical a/c for a house but could be a boon for keeping food or medicine cool in the tropics, without electricity. And design improvements seem possible. Technical fixes for our demand for energy are good but it is also good to be reminded of the cold dark heart of the Universe. Seen against that background, seeing a flower and smiling is a treasure.
Plant science too! It often happens that before we can do an experiment, we have to do another experiment, or two, or many more. I described before that to explain the stable velocity profile, free movement of small molecules through cell-to-cell channels called plasmodesmata might matter. I described last week how I got a line of plants where I could test this. The line drives the expression of a gene that synthesizes a plasmodesmal glue. To be specific, the glue is polysaccharide called callose and the native function of this gene is probably to seal up the channels when the plant gets wounded or invaded by pathogens. Helariutta’s lab (I think!) discovered a mutation in this gene that allows the protein to escape regulation, meaning that in this mutant, there is no need for wounding or other stimuli, the plasmodesmata are always glued shut. This makes for a sick plant. But the line I have uses an inducible promoter to control the gene’s expression: the glue gets made only when the plant sees the inducer, in this case the hormone beta-estradiol. The final complexity is that in addition to inducibility, the gene is turned on only in the endodermis, one of the major tissues in the root.
But the question is: how much inducer should I use? Too little and nothing happens; too much and fundamental developmental processes go awry (other wise known as sick root). I made growth plates with various estradiol concentrations and this past Monday I transplanted one-week old seedlings onto those plates. I had Columbia seedlings (the wild type) that I put on zero and 30 µM estradiol and seedlings of the special line (which I will call pEN7) that went on those concentrations and several in between. During the week, I ticked the position of the root tip, and then on Friday measured the daily growth. The results are here:
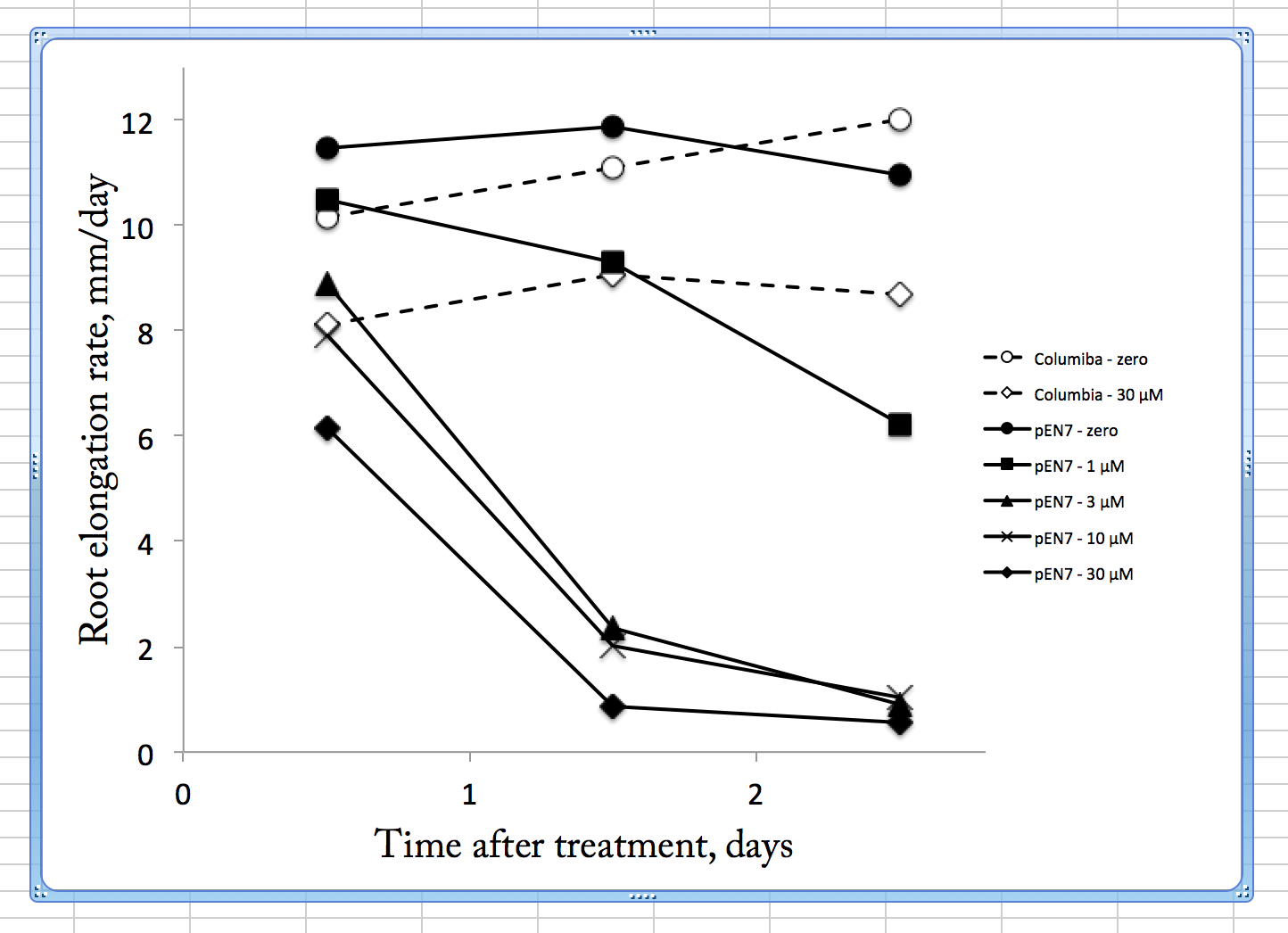
Root growth rate verus time after transfer to the indicated treatment (concentrations refer to beta-estradiol). Symbols are means of three replicate plates, each with 6 seedlings. SEMs are not shown but were generally less than 10% of the mean value.
The figure plots root elongation rate on each day following treatment. In the absence of the inducer (circles), both lines grew about the same rate. The pEN7 line decreased a little in rate on day 3 but this is an artifact, arising because many of the roots had reached the bottom of the plate and hence could not be measured (and those would be the fast growing ones of the population). The wild type on 30 µM estradiol grew more slowly than on zero, indicating that the inducer is not totally inert. Plants do have their own steroid hormones and while the structures differ from those of animals there is overlap and the plant steroid pathways probably account for the growth inhibition here. As for the pEN7 seedlings as exposed to estradiol, the three highest concentrations (30, 10, and 3 µM) behaved similarly, being slightly affected on the first day and crashing on the second. By eye, these seedlings were crooked and had bulbous cells, the latter indicating possible disturbances to root hair development and microtubule cytoskeleton.
Interestingly, on 1 µM the plants were much less affected. While the approximate 50% reduction on day 3 is about the level of growth reduction I am looking for (good sized effect but not too strong), this datum hides variation. Its error bar is much bigger than others because on those plates, at that day, some of the seedlings had crashed (i.e. had really little growth, like the higher concentrations) but others still grew well. That is to say, there might have been NO seedlings growing at that average rate. This suggests that with moderate closure of the plasmodesmata in the endodermis, nothing much happens to growth and as the closure builds up then a developmental switch is thrown and the root crashes. Insofar as mutants whose roots stopped growth were used to identify transcription factors, such as SHORTROOT, that move out of the endodermis through plasmodesmata, this is not that surprising. I was hoping there would be time before such a crash where I could image root growth. But it seems not. The first day’s growth on 30 µM estradiol might offer such a window but as seen from the Columbia, this concentration exerts non-specific effects. I reluctantly conclude that these lines are not going to be useful for my studies.
It happened that Dr Helariutta visited this past week and told me that he has other lines where the plasmodesmata are inducibly closed in other tissues, including cortex and epidermis. He said he would send them and if he does, I’ll see how those work. Stay tuned!