July 26 Experimentation with bells on
In which experiments during my final week at CPIB are described in loving and doubtlessly excessive detail.
Seeing as this past week was the final week of my experimental adventures in CPIB land, I sought to make the most of it. Why should I let little things like cleaning or packing get in the way? I made progress on both root and stem growth and also gave a talk on communicating (writing, speaking, and designing). I will dutifully describe the past week’s lab work here, saving for next week any retrospective (or just plain ol’ retro) musings, assuming that next Sunday won’t see me too dusty and breathless.
First, root growth. I made three more image sequences of wild-type roots growing in agar, thus increasing my total to 12. When I will have time to analyze them, Cthulhu knows, but sitting patiently on the hard disc, they will be waiting. For the very last one of those roots, this Friday, I had gotten about an hour into it when I noticed that something was wrapped around the root, near but not at the tip. It looked like a woolen muffler but was sloughed-off root cap. That I missed it for an hour testifies to my obliviousness. How ?? could I have missed it? The root-cap muffler will be partly stuck in the agar and unlikely to be moving at the same rate as the enshrouded root tip and will thus mess up the resulting data. Part of me wanted to make a massive shrug and settle for 11 roots, but I restrained the shrug to my shoulders, found another root, one free of (visible) anomalies, and captured the sequence.
On the root growth front, there was another development. I had noticed something funny about the times given to the image files. To review: the imaging program (“LabView”) captures a pair of images separated by 60 sec, waits 4 min and captures another pair, continuing for a total of 3 h. The velocity profiles are calculated for the image pairs (based on how much the root moves over the 60 sec). While all this image acquisition is automated (done by LabView), my job during the 3 hours is to move the stage up to keep the root in the field of view. I make a small adjustment every 5 min or so. But if I move the stage during the min when a pair is being acquired, then disaster! This means I have to keep an eye on the times. To do this, I have open on the computer screen the folder (directory) where the image files are being saved and the column with their time of creation is visible. So I get a list of times, that looks like this:
10:01
10:02
10:05
10:06
10:10
10:11
…..
And since the screen also shows the time right now, I can easily tell whether the next acquisition is imminent. All to the good.
But sometimes there was too much time and the minutes would go up by one, so that, for a list like the above, the times would shift to :02, :03 & :11, :12. They never went back. When I first noticed this, some time ago, I dismissed it as rounding error. But eventually I became worried. I asked Darren, and he investigated, discovering that actually the LabView program doesn’t count the time it takes to write the file. That is really weird but it means that, because the images are large and the computer old, the second or two required to write the file are omitted from the time calculation. Thus, instead of every image pair being separated by exactly 60 sec, they are separated by about 60 sec, with some separated by 61 sec (and one of them, in the example I looked at) separated by 62 sec. Likewise, instead of each pair being separated by 4 min, they are separated by 4 min and a bit.
Now, the analysis is not particularly sensitive to the time between pairs, so the difference between 4 min and 4 min, 1 sec is truly inconsequential. However, the time between frames is a different story – the velocity profile is calculated with that time interval in the denominator. Admittedly, one part in 61 is small and unlikely to distort the results. Still, I seek to suppress all forms of noise for these records, and so this timey wimey variation was an unpleasant discovery indeed. Darren wrote a script that pulls out the actual intervals and I will use them, certainly for the new roots, maybe to redo the old ones. I am hopeful that when I set this up at UMass by using Micromanager, instead of LabView, the problem will vanish. But time will tell.
Second, stem growth. The week before last, I did a long segment growth experiment, that was not fully analyzed by last Sunday. Faithful readers will know this because I described the set up last time but no results. Here they are:
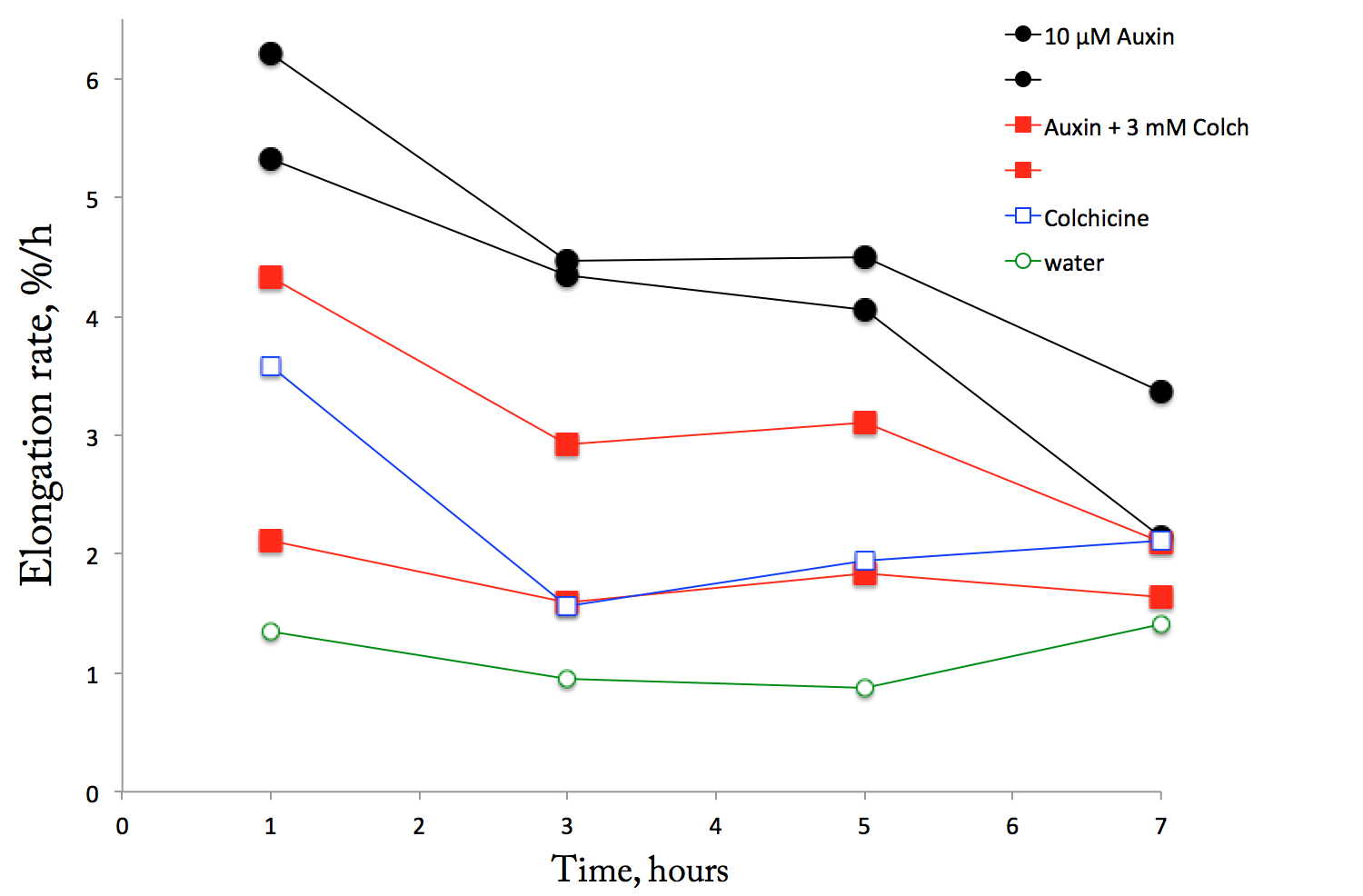
Kinetics of elongation rate for maize mesocotyl segments. Each line is a dish and each dish has nine segments, cut while submerged. Note that auxin (black lines) and auxin + colchicine (red lines) are represented by two dishes each. The standard deviations are not shown because they reach the moon.
Each line represents a dish, containing nine segments; the symbols plot the average rate over a ~2 hour interval. Two of the treatments (auxin, and auxin plus colchicine) had two dishes each; the water and colchicine controls just had one. Yes, this is sub-standard practice, but as this is a pilot to look at the big picture, I can live with that. In oh-so-clear hindsight, I ought to have used three dishes for the main treatments. Still, I learned a few things.
First, I omit the standard deviations because they are big. Wretchedly big. As I have been moaning about for months, the maize mesocotyl segments on auxin grow fast, slow, and everything in between. The averages for the two ‘auxin’ dishes are reasonably close to each other, which is good, but the two ‘auxin + colchicine’ dishes differ by quite a lot. Likewise, the first time point of the colchicine dish (blue) is hilariously high, because of a couple of segments were elongating like crazy.
A novelty of this experiment was using 10 µM IAA (previous ones had used 3 µM). According to plenty of literature there is a rather broad optimum for response to IAA, around 10 µM. I was wondering if giving segments more auxin would help all of them get the message. Apparently not.
Next, on auxin, elongation rate drops by half over the 8 hours. This is typical of results in the literature, though not often characterized. Finally, the colchicine strongly inhibits elongation. In the classic view, this is expected to be concomitant with faster radial expansion rate (“swelling”). There was no hint of swelling. I went out to 8 h to see if I could find swelling after a lag, but evidently this was not long enough. Disappointing results, because I am hoping to use this system to study swelling. It is plausible that the segments respond to the loss of microtubules as a stress (along with whatever is causing the general decline of response to auxin), shut down operations. I saw a similar large drop in elongation rate some months ago when I used a different microtubule inhibitor (oryzalin). It might be possible to stimulate swelling with less extreme doses, or by using cytokinin, which in the literature has also been reported to make segments get fat. Something for the table at UMass.
And one more thing. Last week’s experiment! It was simple. I used two dishes with tomato hypocotyls because in this species there are cuticle mutants that might help with the problem of drug delivery. The tomato hypocotyls are thin and fussing about with them in the dark with my Cyclops eye was something I hope I never need to do again. Many of them were not cut clean through and probably damaged with the tweezers. Still, they managed to respond to auxin more or less like the other species. I expect I can find a way to be gentler if I can work under dim red light, of which I live in hopes for many reasons.
And last, maybe not least, the remaining four dishes. Back to maize and the question of the decline in growth rate. I tried adding 5 mM glucose to the dishes. In an earlier experiment, I found that 50 mM sucrose was a downer, but that’s ten times higher. Maybe the decline is caused by running out of juice? Nope, not according to the results last week. I had two dishes for each treatment, and looked at growth from zero to three hours, and from three to six hours. Comparing the first interval to the second, both the ‘auxin’ and the ‘auxin + glucose’ segments declined similarly. However, interestingly, the rate of elongation was distinctly higher with glucose. That’s not a bad thing. Further exploration required.
Yes, well, further exploration is always required. But, is it worth it. I think so, but I will write about that next week as I attempt to summarize and encapsulate the year. I wonder…