November 23: Out-reaching for the skies
How do oats, peas, beans, and barely grow? In which I describe my efforts to give a Knowledge Transfer Seminar to enlighten the local non-academics about life in the Lab-Fab universe.
Fresh from the thrill of explaining to school kids why one would give a root a CTscan, you might think I would be dead eager to return to the calmer world of pipettes and test tubes. And you would be correct. But life. Not long after my arrival in these cloudy climes I was invited to give a Knowledge Transfer Seminar, a rather grandly titled affair that is actually meant to provide a venue for academics to explain to the local non-academic staff what it is they do. This venue has been going for some years at Sutton Bonington with popularity and even draws a handful of townsfolk curious to know what is going on within the mysterious gates of campus.
As fate would have it, the date in question was this past Thursday. I gave a seminar on the mechanism of plant growth, entitled “How do oats, peas, beans, and barley grow?” I decided to explain from first principles how a plant cell enlarges. I talked about the pivotal position of growth in the plant’s life, not just ‘to get bigger’ as in animals, which is of course important, but also to acclimate to the environment, where a plant grows new organs throughout its life with sizes and shapes tailored to what is going on around it. I then introduced the concepts of an elastic material and of water chemical potential energy and used them to explain the force balance that allows a non-woody plant stands up. The low water chemical potential inside the cell drives water uptake which stretches the elastic cell wall, which as a result of being stretched exerts a force to oppose water entry. This mechanical equilibrium keeps (non-woody) plants lifted up off the ground.
With that established, to deal with a growing cell, I then added the concept of a plastic element, the delightfully named “dashpot”. In a growing cell, the elastic components are linked to plastic ones, which allow growth to occur by stress-relaxation. That is, the stretched strings pull out the dashpots and thus shorten, exert a lower force, and consequently water enters to re-stretch the springs. Voilá! growth! (see figure below)
Finally, I turned to the question of growth direction and introduced the audience to the delights of cellulose microfibrils and cortical microtubules. This gets into the special complexity of biology but was worth it to show some pretty pictures of microtubules, which are beautiful, and a nice diagram of cellulose, which is one of nature’s wonders. I also talked about stresses in the cell and illustrated the interplay of stress (force) and compliance (growth).
This was challenging to do in a way that non specialists could follow and that is why I spent most of the week planning what I would say exactly and making illustrations. Here is one example:
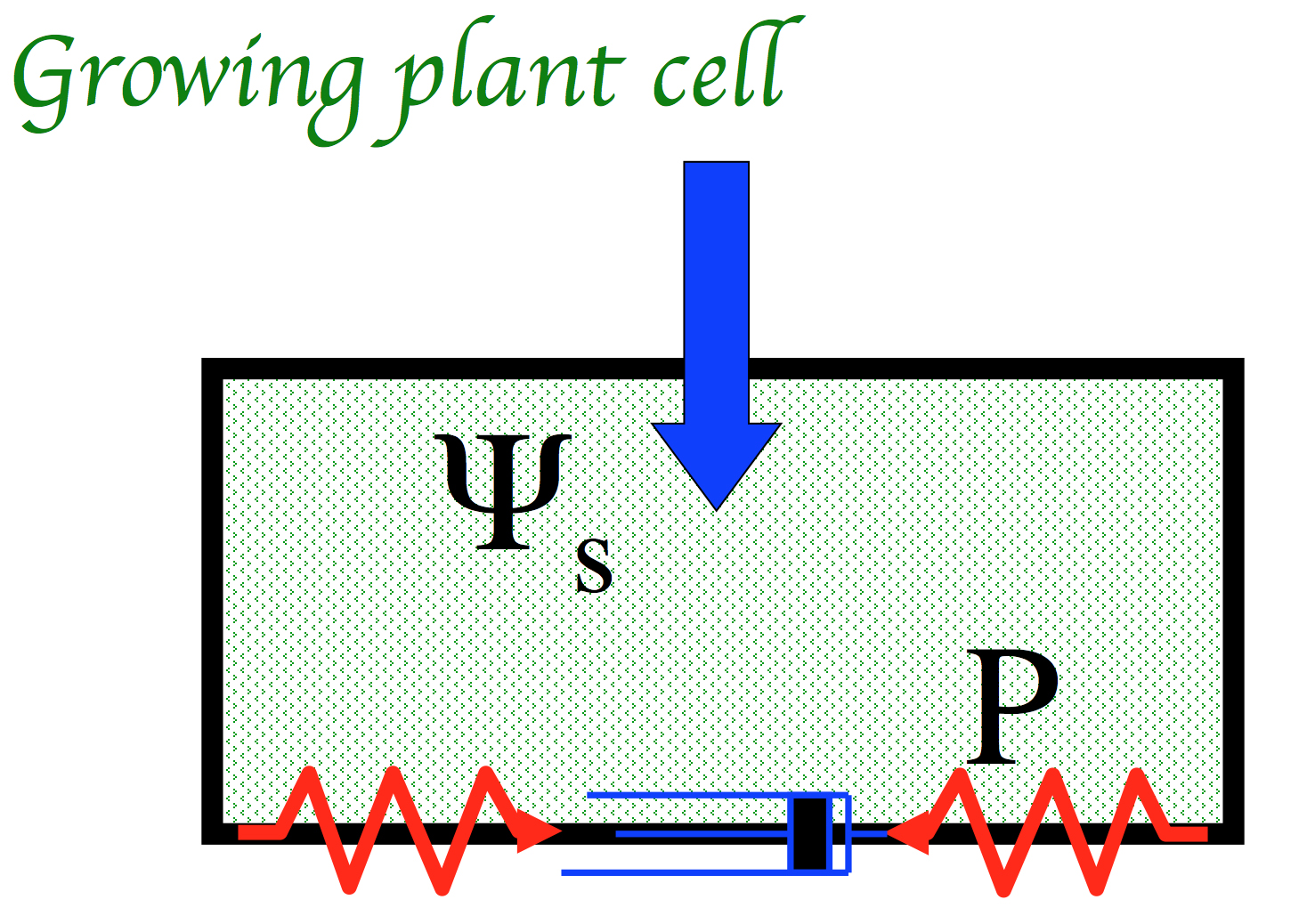
Components underlying plant growth. The cell wall contains elastic (springs) and plastic (dashpot) elements. Water moving in from high potential to low stretches the springs, generating turgor (P) which does work against the dashpots. This relaxes the springs and allows more water entry.
I like illustrations large and clear and these are more difficult to hoak up than one might think. Also I had never given this talk before so while I had bits and pieces from other lectures, quite a few slides needed to be made from scratch or carefully edited.
Based on the feedback, I think the audience stayed awake and learned a thing or two about growth. We do know something about how oats, peas, beans, and barely grow, even if there remain many things we don’t know. That after all is why I stay in the laboratory.
Still, I managed to do a little bit of work. I had planned to make refinements on the segment imaging workstation but the plants I sent up on the Monday did not grow well enough, so I put that off for this coming week. But I did get to make a set of Petri plates containing growth medium into which I mixed beta-estradiol at various concentrations. No, I am not interested in changing the sex of arabidopsis, which of course would not respond to animal hormones. Rather, I made the plates to induce a gene in a special line of plants. I got the line from my former PhD student, Shuang Wu, who is now working with Kim Gallagher at Penn. The line contain a gene that will occlude plasmodesmata, those channels that allow direct passage between cells of molecules lighter than about 1 kDa (that is smaller than nearly all proteins and RNAs but larger than typical metabolites and signal molecules). Now, occluding plasmodesmata on a permanent basis would lead to a dead or deeply disturbed plant so Xiao Wu (as I like to call him in the Chinese way) hooked up a promoter to the gene so it will only turn on in the presence of estradiol. And because estradiol has no natural business in plants, turning on this gene is all that hormone should do.
Xiao Wu told me that 20 µM estradiol would induce the gene massively, so I prepared plates without any estradiol and containing 1, 3, 10, and 30 µM estradiol. On Monday I will transplant seedlings onto the plates and tick the roots (as described here 19/10 slowly ramping up) for a few days. I will have wild-type seedlings on zero and 30 µM estradiol from which I can tell whether the hormone really is inert and the special line will be on all of the concentrations. What I would like to find would be a concentration that reduces growth but doesn’t stop it because while stopped growth would be dramatic, it would give me nowt to study. But for slower growth, I can ask whether, along with the lowered rate, the stability of the velocity profile is altered. This will test to what extent stabilization of the velocity profile requires transport through plasmodesmata. There are reasons to think it might, and other reasons to think it might not. So when results are in hand, I might get into those.
An important issue is that Xiao Wu set up this genetic light and magic show to happen specifically in a single tissue, the endodermis. It is not clear that the endodermis would be privileged in regard to controlling growth but it might be. Xiao Wu tells me that other lines are in progress, so perhaps eventually they can be checked too.